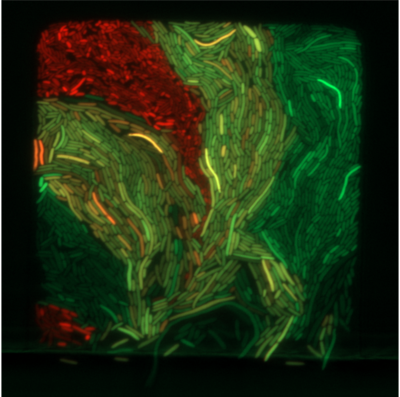
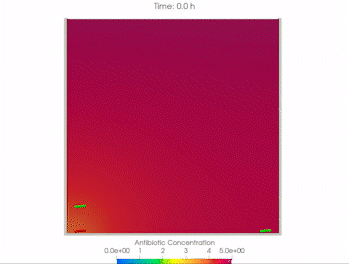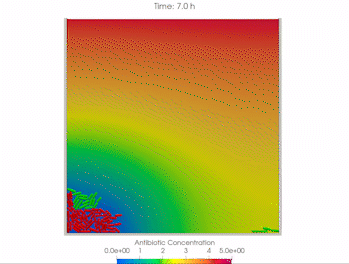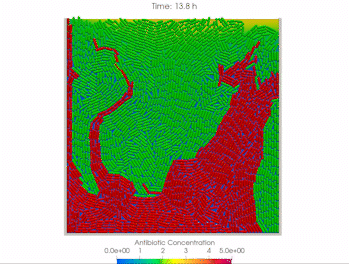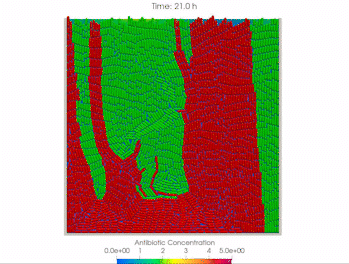
Welcome to the Ingalls Quantitative Cell Biology Lab in the Department of Applied Mathematics at the University of Waterloo.
We use mathematical models and experimental methods to investigate the behaviour of intracellular molecular networks and cell-to-cell interactions. This work ranges from fundamental studies of biology to applications in biotechnology and health.